J Cell Mol Med. 2013 Aug; 17(8): 958–965.
Published online 2013 Jun 26. doi: 10.1111/jcmm.12088
Martin L Pall*
Author information ► Article notes ► Copyright and License information ►
This article has been cited by other articles in PMC.
Go to:
Abstract
The direct targets of extremely low and microwave frequency range electromagnetic fields (EMFs) in producing non-thermal effects have not been clearly established. However, studies in the literature, reviewed here, provide substantial support for such direct targets. Twenty-three studies have shown that voltage-gated calcium channels (VGCCs) produce these and other EMF effects, such that the L-type or other VGCC blockers block or greatly lower diverse EMF effects. Furthermore, the voltage-gated properties of these channels may provide biophysically plausible mechanisms for EMF biological effects. Downstream responses of such EMF exposures may be mediated through Ca2+/calmodulin stimulation of nitric oxide synthesis. Potentially, physiological/therapeutic responses may be largely as a result of nitric oxide-cGMP-protein kinase G pathway stimulation. A well-studied example of such an apparent therapeutic response, EMF stimulation of bone growth, appears to work along this pathway. However, pathophysiological responses to EMFs may be as a result of nitric oxide-peroxynitrite-oxidative stress pathway of action. A single such well-documented example, EMF induction of DNA single-strand breaks in cells, as measured by alkaline comet assays, is reviewed here. Such single-strand breaks are known to be produced through the action of this pathway. Data on the mechanism of EMF induction of such breaks are limited; what data are available support this proposed mechanism. Other Ca2+-mediated regulatory changes, independent of nitric oxide, may also have roles. This article reviews, then, a substantially supported set of targets, VGCCs, whose stimulation produces non-thermal EMF responses by humans/higher animals with downstream effects involving Ca2+/calmodulin-dependent nitric oxide increases, which may explain therapeutic and pathophysiological effects.
Keywords: intracellular Ca2+, voltage-gated calcium channels, low frequency electromagnetic field exposure, nitric oxide, oxidative stress, calcium channel blockers
Introduction
An understanding of the complex biology of the effects of electromagnetic fields (EMFs) on human/higher animal biology inevitably must be derived from an understanding of the target or targets of such fields in the impacted cells and tissues. Despite this, no understanding has been forthcoming on what those targets are and how they may lead to the complex biological responses to EMFs composed of low-energy photons. The great puzzle, here, is that these EMFs are comprised of low-energy photons, those with insufficient energy to individually influence the chemistry of the cell, raising the question of how non-thermal effects of such EMFs can possibly occur. The author has found that there is a substantial literature possibly pointing to the direct targets of such EMFs and it is the goal of this study to review that evidence as well as review how those targets may lead to the complex biology of EMF exposure.
The role of increased intracellular Ca2+ following EMF exposure was already well documented more than 20 years ago, when Walleczek [1] reviewed the role of changes in calcium signalling that were produced in response EMF exposures. Other, more recent studies have confirmed the role of increased intracellular Ca2+following EMF exposure, a few of which are discussed below. His review [1] included two studies [2, 3] that showed that the L-type voltage-gated channel blocker, verapamil could lower or block changes in response to EMFs. The properties of voltage-gated calcium channels (VGCCs) have been reviewed elsewhere [4]. Subsequently, extensive evidence has been published clearly showing that the EMF exposure can act to produce excessive activity of the VGCCs in many cell types [5–26] suggesting that these may be direct targets of EMF exposure. Many of these studies implicate specifically the L-type VGCCs such that various L-type calcium channel blockers can block responses to EMF exposure (Table 1). However, other studies have shown lowered responses produced by other types of calcium channel blockers including N-type, P/Q-type, and T-type blockers (Table 1), showing that other VGCCs may have important roles. Diverse responses to EMFs are reported to be blocked by such calcium channel blockers (Table 1), suggesting that most if not all EMF-mediated responses may be produced through VGCC stimulation. Voltage-gated calcium channels are essential to the responses produced by extremely low frequency (including 50/60 Hz) EMFs and also to microwave frequency range EMFs, nanosecond EMF pulses, and static electrical and magnetic fields (Table 1).
Published online 2013 Jun 26. doi: 10.1111/jcmm.12088
Martin L Pall*
Author information ► Article notes ► Copyright and License information ►
This article has been cited by other articles in PMC.
Go to:
Abstract
The direct targets of extremely low and microwave frequency range electromagnetic fields (EMFs) in producing non-thermal effects have not been clearly established. However, studies in the literature, reviewed here, provide substantial support for such direct targets. Twenty-three studies have shown that voltage-gated calcium channels (VGCCs) produce these and other EMF effects, such that the L-type or other VGCC blockers block or greatly lower diverse EMF effects. Furthermore, the voltage-gated properties of these channels may provide biophysically plausible mechanisms for EMF biological effects. Downstream responses of such EMF exposures may be mediated through Ca2+/calmodulin stimulation of nitric oxide synthesis. Potentially, physiological/therapeutic responses may be largely as a result of nitric oxide-cGMP-protein kinase G pathway stimulation. A well-studied example of such an apparent therapeutic response, EMF stimulation of bone growth, appears to work along this pathway. However, pathophysiological responses to EMFs may be as a result of nitric oxide-peroxynitrite-oxidative stress pathway of action. A single such well-documented example, EMF induction of DNA single-strand breaks in cells, as measured by alkaline comet assays, is reviewed here. Such single-strand breaks are known to be produced through the action of this pathway. Data on the mechanism of EMF induction of such breaks are limited; what data are available support this proposed mechanism. Other Ca2+-mediated regulatory changes, independent of nitric oxide, may also have roles. This article reviews, then, a substantially supported set of targets, VGCCs, whose stimulation produces non-thermal EMF responses by humans/higher animals with downstream effects involving Ca2+/calmodulin-dependent nitric oxide increases, which may explain therapeutic and pathophysiological effects.
Keywords: intracellular Ca2+, voltage-gated calcium channels, low frequency electromagnetic field exposure, nitric oxide, oxidative stress, calcium channel blockers
- Introduction
- Possible modes of action following voltage-gated calcium channel stimulation
- Therapeutic bone-growth stimulation via Ca2+/nitric oxide/cGMP/protein kinase G
- Ca2+/nitric oxide/peroxynitrite and pathophysiological responses to EMF exposures: the example of single-strand DNA breaks
- Discussion and conclusions
Introduction
An understanding of the complex biology of the effects of electromagnetic fields (EMFs) on human/higher animal biology inevitably must be derived from an understanding of the target or targets of such fields in the impacted cells and tissues. Despite this, no understanding has been forthcoming on what those targets are and how they may lead to the complex biological responses to EMFs composed of low-energy photons. The great puzzle, here, is that these EMFs are comprised of low-energy photons, those with insufficient energy to individually influence the chemistry of the cell, raising the question of how non-thermal effects of such EMFs can possibly occur. The author has found that there is a substantial literature possibly pointing to the direct targets of such EMFs and it is the goal of this study to review that evidence as well as review how those targets may lead to the complex biology of EMF exposure.
The role of increased intracellular Ca2+ following EMF exposure was already well documented more than 20 years ago, when Walleczek [1] reviewed the role of changes in calcium signalling that were produced in response EMF exposures. Other, more recent studies have confirmed the role of increased intracellular Ca2+following EMF exposure, a few of which are discussed below. His review [1] included two studies [2, 3] that showed that the L-type voltage-gated channel blocker, verapamil could lower or block changes in response to EMFs. The properties of voltage-gated calcium channels (VGCCs) have been reviewed elsewhere [4]. Subsequently, extensive evidence has been published clearly showing that the EMF exposure can act to produce excessive activity of the VGCCs in many cell types [5–26] suggesting that these may be direct targets of EMF exposure. Many of these studies implicate specifically the L-type VGCCs such that various L-type calcium channel blockers can block responses to EMF exposure (Table 1). However, other studies have shown lowered responses produced by other types of calcium channel blockers including N-type, P/Q-type, and T-type blockers (Table 1), showing that other VGCCs may have important roles. Diverse responses to EMFs are reported to be blocked by such calcium channel blockers (Table 1), suggesting that most if not all EMF-mediated responses may be produced through VGCC stimulation. Voltage-gated calcium channels are essential to the responses produced by extremely low frequency (including 50/60 Hz) EMFs and also to microwave frequency range EMFs, nanosecond EMF pulses, and static electrical and magnetic fields (Table 1).
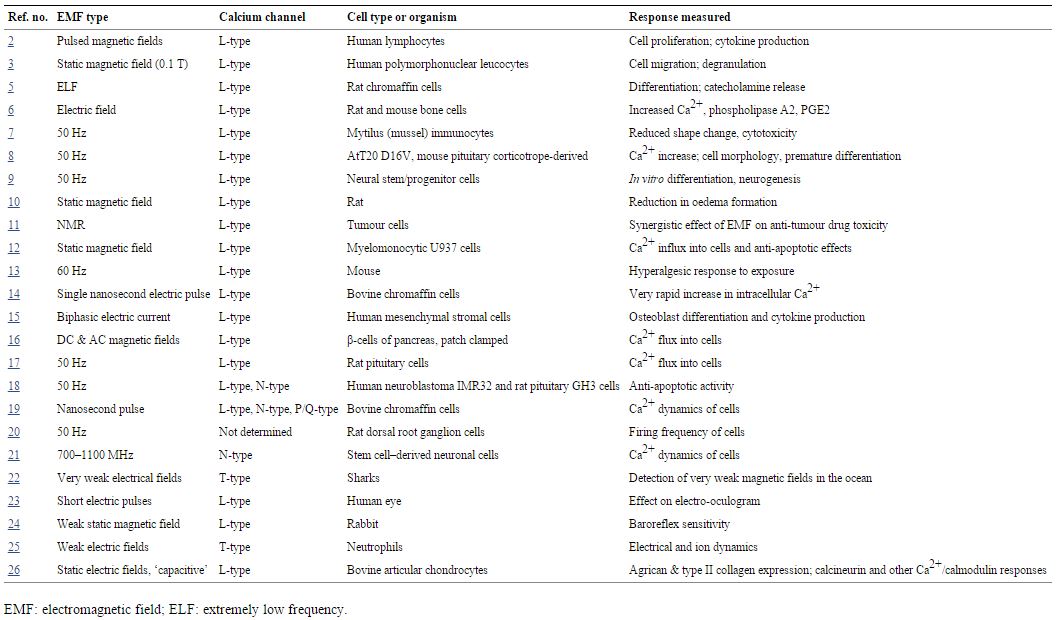
Table 1
EMF responses blocked or lowered by calcium channel blockers
In a recent study, Pilla [27] showed that an increase in intracellular Ca2+ must have occurred almost immediately after EMF exposure, producing a Ca2+/calmodulin-dependent increase in nitric oxide occurring in less than 5 sec. Although Pilla [27] did not test whether VGCC stimulation was involved in his study, there are few alternatives that can produce such a rapid Ca2+ response, none of which has been implicated in EMF responses. Other studies, each involving VGCCs, summarized in Table 1, also showed rapid Ca2+increases following EMF exposure [8, 16, 17, 19, 21]. The rapidity of these responses rule out many types of regulatory interactions as being involved in producing the increased VGCC activity following EMF exposure and suggests, therefore, that VGCC stimulation in the plasma membrane is directly produced by EMF exposure.
Go to:
Possible modes of action following VGCC stimulation
The increased intracellular Ca2+ produced by such VGCC activation may lead to multiple regulatory responses, including the increased nitric oxide levels produced through the action of the two Ca2+/calmodulin-dependent nitric oxide synthases, nNOS and eNOS. Increased nitric oxide levels typically act in a physiological context through increased synthesis of cGMP and subsequent activation of protein kinase G [28, 29]. In contrast, in most pathophysiological contexts, nitric oxide reacts with superoxide to form peroxynitrite, a potent non-radical oxidant [30, 31], which can produce radical products, including hydroxyl radical and NO2 radical [32].
Go to:
Therapeutic bone-growth stimulation via Ca2+/nitric oxide/cGMP/protein kinase G
An example of a therapeutic effect for bone repair of EMF exposure in various medical situations includes increasing osteoblast differentiation and maturation and has been reviewed repeatedly [33–44]. The effects of EMF exposure on bone cannot be challenged, although there is still considerable question about the best ways to apply this clinically [33–44]. Our focus, here, is to consider possible mechanisms of action. Multiple studies have implicated increased Ca2+ and nitric oxide in the EMF stimulation of bone growth [44–49]; three have also implicated increased cGMP and protein kinase G activity [46, 48, 49]. In addition, studies on other regulatory stimuli leading to increased bone growth have also implicated increased cGMP levels and protein kinase G in this response [50–56]. In summary, then, it can be seen from the above that there is a very well-documented action of EMFs in stimulating osteoblasts and bone growth. The available data, although limited, support the action of the main pathway involved in physiological responses to Ca2+ and nitric oxide, namely Ca2+/nitric oxide/cGMP/protein kinase G in producing such stimulation.
Go to:
Ca2+/nitric oxide/peroxynitrite and pathophysiological responses to EMF exposures: the example of single-strand DNA breaks
As was noted above, most of the pathophysiological effects of nitric oxide are mediated through peroxynitrite elevation and consequent oxidative stress. There are many reviews and other studies, implicating oxidative stress in generating pathophysiological effects of EMF exposure [see for example [57–64]]. In some of these studies, the rise in oxidative stress markers parallels the rise in nitric oxide, suggesting a peroxynitrite-mediated mechanism [64–67].
Peroxynitrite elevation is usually measured through a marker of peroxynitrite-mediated protein nitration, 3-nitrotyrosine (3-NT). There are four studies where 3-NT levels were measured before and after EMF exposure [66, 68–70]. Each of these studies provides some evidence supporting the view that EMF exposure increases levels of peroxynitrite and therefore 3-NT levels [66, 68–70]. Although these cannot be taken as definitive, when considered along with the evidence on oxidative stress and elevated nitric oxide production in response to EMF exposure, they strongly suggest a peroxynitrite-mediated mechanism of oxidative stress in response to EMFs.
Such a peroxynitrite-mediated mechanism may explain the many studies showing the single-stranded breaks in DNA, as shown by alkaline comet assays or the similar microgel electrophoresis assay, following EMF exposures in most such studies [71–89], but not in all [90–97]. Some of the factors that are reported to influence whether such DNA single-strand breaks are detected after EMF exposure include the type of cell studied [79, 86], dosage of EMF exposure [78] and the type of EMF exposure studied [73, 77]. Oxidative stress and free radicals have roles, both because there is a concomitant increase in oxidative stress and because antioxidants have been shown to greatly lower the generation of DNA single-strand breaks following EMF exposure [72, 75, 81, 82] as has also been shown for peroxynitrite-mediated DNA breaks produced under other conditions. It has also been shown that one can block the generation of DNA single-strand breaks with a nitric oxide synthase inhibitors [82].
Peroxynitrite has been shown to produce single-strand DNA breaks [98–100], a process that is inhibited by many but not all antioxidants [99, 100]. It can be seen from this that the data on generation of single-strand DNA breaks, although quite limited, support a mechanism involving nitric oxide/peroxynitrite/free radical (oxidative stress). Although the data on the possible role of peroxynitrite in EMF-induced DNA single-strand breaks are limited, what data are available supports such a peroxynitrite role.
Go to:
Discussion and conclusions
How do EMFs composed of low-energy photons produce non-thermal biological changes, both pathophysiological and, in some cases, potentially therapeutic, in humans and higher animals? It may be surprising that the answer to this question has been hiding in plain sight in the scientific literature. However, in this era of highly focused and highly specialized science, few of us have the time to read the relevant literature, let alone organize the information found within it in useful and critical ways.
This study shows that:
The possible role of peroxynitrite as opposed to protein kinase G in producing pathophysiological responses to EMF exposure raises the question of whether there are practical approaches to avoiding such responses? Typically peroxynitrite levels can be highly elevated when both of its precursors, nitric oxide and superoxide, are high. Consequently, agents that lower nitric oxide synthase activity and agents that raise superoxide dismutases (SODs, the enzymes that degrade superoxide) such as phenolics and other Nrf2 activators that induce SOD activity [101], as well as calcium channel blockers may be useful. Having said that, this is a complex area, where other approaches should be considered, as well.
Although the various EMF exposures as well as static electrical field exposures can act to change the electrical voltage-gradient across the plasma membrane and may, therefore, be expected to stimulate VGCCs through their voltage-gated properties, it may be surprising that static magnetic fields also act to activate VGCCs because static magnetic fields do not induce electrical changes on static objects. However, cells are far from static. Such phenomena as cell ruffling [102],[103] may be relevant, where thin cytoplasmic sheets bounded on both sides by plasma membrane move rapidly. Such rapid movement of the electrically conducting cytoplasm, may be expected to influence the electrical charge across the plasma membrane, thus potentially stimulating the VGCCs.
Earlier modelling of electrical effects across plasma membranes of EMF exposures suggested that such electrical effects were likely to be too small to explain EMF effects at levels reported to produce biological changes (see, for example [22]). However, more recent and presumably more biologically plausible modelling have suggested that such electrical effects may be much more substantial [104–109] and may, therefore, act to directly stimulate VGCCs.
Direct stimulation of VGCCs by partial depolarization across the plasma membrane is suggested by the following observations discussed in this review:
The only detailed alternative to the mechanism of non-thermal EMF effects discussed here, to my knowledge, is the hypothesis of Friedman et al. [114] and supported by Desai et al. [115] where the apparent initial response to EMF exposure was proposed to be NADH oxidase activation, leading to oxidative stress and downstream regulatory effects. Although they provide some correlative evidence for a possible role of NADH oxidase [114], the only causal evidence is based on a presumed specific inhibitor of NADH oxidase, diphenyleneiodonium (DPI). However, DPI has been shown to be a non-specific cation channel blocker [116], clearly showing a lack of such specificity and suggesting that it may act, in part, as a VGCC blocker. Consequently, a causal role for NADH oxidase in responses to EMF exposure must be considered to be undocumented.
In summary, the non-thermal actions of EMFs composed of low-energy photons have been a great puzzle, because such photons are insufficiently energetic to directly influence the chemistry of cells. The current review provides support for a pathway of the biological action of ultralow frequency and microwave EMFs, nanosecond pulses and static electrical or magnetic fields: EMF activation of VGCCs leads to rapid elevation of intracellular Ca2+, nitric oxide and in some cases at least, peroxynitrite. Potentially therapeutic effects may be mediated through the Ca2+/nitric oxide/cGMP/protein kinase G pathway. Pathophysiological effects may be mediated through the Ca2+/nitric oxide/peroxynitrite pathway. Other Ca2+-mediated effects may have roles as well, as suggested by Xu et al. [26].
Go to:
Conflicts of interest
The author confirms that there are no conflicts of interest.
Go to:
References
1. Walleczek J. Electromagnetic field effects on cells of the immune system: the role of calcium signaling.FASEB J. 1992;6:3177–85. [PubMed]
2. Cadossi R, Emilia G, Ceccherelli G. 1988 Lymphocytes and pulsing magnetic fields. In: Marino EE, et al., editors. Modern bioelectricity. New York: Dekker; 1998. pp. 451–96.
3. Papatheofanis FJ. Use of calcium channel antagonists as magnetoprotective agents. Radiat Res.1990;122:24–8. [PubMed]
4. Catterall WA. Structure and regulation of voltage-gated Ca2+ channels. Annu Rev Cell Dev Biol.2000;16:521–55. [PubMed]
5. Morgado-Valle C, Verdugo-Díaz L, García DE, et al. The role of voltage-gated Ca2+ channels in neurite growth of cultured chromaffin cells induced by extremely low frequency (ELF) magnetic field stimulation.Cell Tissue Res. 1998;291:217–30. [PubMed]
6. Lorich DG, Brighton CT, Gupta R, et al. Biochemical pathway mediating the response of bone cells to capacitive coupling. Clin Orthop Relat Res. 1998:246–56. [PubMed]
7. Gobba F, Malagoli D, Ottaviani E. Effects of 50 Hz magnetic fields on fMLP-induced shape changes in invertebrate immunocytes: the role of calcium ion channels. Bioelectromagnetics. 2003;24:277–82.[PubMed]
8. Lisi A, Ledda M, Rosola E, et al. Extremely low frequency electromagnetic field exposure promotes differentiation of pituitary corticotrope-derived AtT20 D16V cells. Bioelectromagnetics. 2006;27:641–51.[PubMed]
9. Piacentini R, Ripoli C, Mezzogori D, et al. Extremely low-frequency electromagnetic fields promote in vitro neurogenesis via upregulation of Ca(v)1-channel activity. J Cell Physiol. 2008;215:129–39. [PubMed]
10. Morris CE, Skalak TC. Acute exposure to a moderate strength static magnetic field reduces edema formation in rats. Am J Physiol Heart Circ Physiol. 2008;294:H50–7. [PubMed]
11. Ghibelli L, Cerella C, Cordisco S, et al. NMR exposure sensitizes tumor cells to apoptosis. Apoptosis.2006;11:359–65. [PubMed]
12. Fanelli C, Coppola S, Barone R, et al. Magnetic fields increase cell survival by inhibiting apoptosis viamodulation of Ca2+ influx. FASEB J. 1999;13:95–102. [PubMed]
13. Jeong JH, Kum C, Choi HJ, et al. Extremely low frequency magnetic field induces hyperalgesia in mice modulated by nitric oxide synthesis. Life Sci. 2006;78:1407–12. [PubMed]
14. Vernier PT, Sun Y, Chen MT, et al. Nanosecond electric pulse-induced calcium entry into chromaffin cells. Bioelectrochemistry. 2008;73:1–4. [PubMed]
15. Kim IS, Song JK, Song YM, et al. Novel effect of biphasic electric current on in vitro osteogenesis and cytokine production in human mesenchymal stromal cells. Tissue Eng Part A. 2009;15:2411–22. [PubMed]
16. Höjevik P, Sandblom J, Galt S, et al. Ca2+ ion transport through patch-clamped cells exposed to magnetic fields. Bioelectromagnetics. 1995;16:33–40. [PubMed]
17. Barbier E, Vetret B, Dufy B. Stimulation of Ca2+ influx in rat pituitary cells under exposure to a 50 Hz magnetic field. Bioelectromagnetics. 1996;17:303–11. [PubMed]
18. Grassi C, D'Ascenzo M, Torsello A, et al. Effects of 50 Hz electromagnetic fields on voltage-gated Ca2+channels and their role in modulation of neuroendocrine cell proliferation and death. Cell Calcium.2004;35:307–15. [PubMed]
19. Craviso GL, Choe S, Chatterjee P, et al. Nanosecond electric pulses: a novel stimulus for triggering Ca2+influx into chromaffin cells via voltage-gated Ca2+ channels. Cell Mol Neurobiol. 2010;30:1259–65.[PubMed]
20. Marchionni I, Paffi A, Pellegrino M, et al. Comparison between low-level 50 Hz and 900 MHz electromagnetic stimulation on single channel ionic currents and on firing frequency in dorsal root ganglion isolated neurons. Biochim Biophys Acta. 2006;1758:597–605. [PubMed]
21. Rao VS, Titushkin IA, Moros EG, et al. Nonthermal effects of radiofrequency-field exposure on calcium dynamics in stem cell-derived neuronal cells: elucidation of calcium pathways. Radiat Res. 2008;169:319–29. [PubMed]
22. Adair RK, Astumian RD, Weaver JC. Detection of weak electric fields by sharks, rays and skates.Chaos. 1998;8:576–87. [PubMed]
23. Constable PA. Nifedipine alters the light-rise of the electro-oculogram in man. Fraefes Arch Clin Exp Ophthalmol. 2011;249:677–84. [PubMed]
24. Gmitrov J, Ohkuba C. Verapamil protective effect on natural and artificial magnetic field cardiovascular impact. Bioelectromagnetics. 2002;23:531–41. [PubMed]
25. Kindzelskii AL, Petty HR. Ion channel clustering enhances weak electric field detection by neutrophils: apparent role of SKF96365-sensitive cation channels and myeloperoxidase trafficking cellular responses. Eur Biophys J. 2005;35:1–26. [PubMed]
26. Xu J, Wang W, Clark CC, et al. Signal transduction in electrically stimulated articular chondrocytes involves translocation of extracellular calcium through voltage-gated channels. Osteoarthritis Cartilage.2009;17:397–405. [PubMed]
27. Pilla AA. Electromagnetic fields instantaneously modulate nitric oxide signaling in challenged biological systems. Biochem Biophys Res Commun. 2012;426:330–3. [PubMed]
28. McDonald LJ, Murad F. Nitric oxide and cyclic GMP signaling. Proc Soc Exp Biol Med. 1996;211:1–6.[PubMed]
29. Francis SH, Busch JL, Corbin JD, et al. cGMP-dependent protein kinases and cGMP phosphodiesterases in nitric oxide and cGMP action. Pharmacol Rev. 2010;62:525–63. [PMC free article] [PubMed]
30. Pacher P, Beckman JS, Liaudet L. Nitric oxide and peroxynitrite in health and disease. Physiol Rev.2007;87:315–424. [PMC free article] [PubMed]
31. Pryor WA, Squadrito GL. The chemistry of peroxynitrite: a product from the reaction of nitric oxide with superoxide. Am J Physiol. 1995;268:L699–722. [PubMed]
32. Lymar SV, Khairutdinov RF, Hurst JK. Hydroxyl radical formation by O-O bond homolysis in peroxynitrous acid. Inorg Chem. 2003;42:5259–66. [PubMed]
33. Ryabi JT. Clinical effects of electromagnetic fields on fracture healing. Clin Orthop Relat Res.1998;355(Suppl. l):S205–15. [PubMed]
34. Oishi M, Onesti ST. Electrical bone graft stimulation for spinal fusion: a review. Neurosurgery.2000;47:1041–55. [PubMed]
35. Aaron RK, Ciombor DM, Simon BJ. Treatment of nonunions with electric and electromagnetic fields.Clin Orthop Relat Res. 2004;10:579–93. [PubMed]
36. Goldstein C, Sprague S, Petrisor BA. Electrical stimulation for fracture healing: current evidence. J Orthop Trauma. 2010;24(Suppl. 1):S62–5. [PubMed]
37. Demitriou R, Babis GC. Biomaterial osseointegration enhancement with biophysical stimulation. J Musculoskelet Neuronal Interact. 2007;7:253–65. [PubMed]
38. Griffin XL, Warner F, Costa M. The role of electromagnetic stimulation in the management of established non-union lf long bone fractures: what is the evidence? Injury. 2008;39:419–29. [PubMed]
39. Huang LQ, He HC, He CQ, et al. Clinical update of pulsed electromagnetic fields on osteroporosis. Chin Med J. 2008;121:2095–9. [PubMed]
40. Groah SL, Lichy AM, Libin AV, et al. Intensive electrical stimulation attenuates femoral bone loss in acute spinal cord injury. PM R. 2010;2:1080–7. [PubMed]
41. Schidt-Rohlfing B, Silny J, Gavenis K, et al. Electromagnetic fields, electric current and bone healing – what is the evidence? Z Orthop Unfall. 2011;149:265–70. [PubMed]
42. Griffin XL, Costa ML, Parsons N, et al. Electromagnetic field stimulation for treating delayed union or non-union of long bone fractures in adults. Cochrane Database Syst Rev. 2011:CDO08471. doi:10.1002/14651858.CD008471.pub2. [PubMed]
43. Chalidis B, Sachinis N, Assiotis A, et al. Stimulation of bone formation and fracture healing with pulsed electromagnetic fields: biologic responses and clinical implications. Int J Immunopathol Pharmacol.2011;24(1 Suppl. 2):17020. [PubMed]
44. Zhong C, Zhao TF, Xu ZJ, et al. Effects of electromagnetic fields on bone regeneration in experimental and clinical studies: a review of the literature. Chin Med J. 2012;125:367–72. [PubMed]
45. Diniz P, Soejima K, Ito G. Nitric oxide mediates the effects of pulsed electromagnetic field stimulation on the osteoblast proliferation and differentiation. Nitric Oxide. 2002;7:18–23. [PubMed]
46. Fitzsimmons RJ, Gordon SL, Ganey T, et al. A pulsing electric field (PEF) increases human chondrocyte proliferation through a transduction pathway involving nitric oxide signaling. J Orthopaedic Res.2008;26:854–9. [PubMed]
47. Lin H-Y, Lin Y-J. In vitro effects of low frequency electromagnetic fields on osteoblast proliferation and maturation in an inflammatory environment. Bioelectromagnetics. 2011;32:552–60. [PubMed]
48. Cheng G, Zhai Y, Chen K, et al. Sinusoidal electromagnetic field stimulates rat osteoblast differentiation and maturation via activation of NO-cGMP-PKG pathway. Nitric Oxide. 2011;25:316–25. [PubMed]
49. Pilla A, Fitzsimmons R, Muehsam D, et al. Electromagnetic fields as first messenger in biological signaling: application to calmodulin-dependent signaling in tissue repair. Biochim Biophys Acta.2011;1810:1236–45. [PubMed]
50. Rangaswami H, Schwappacher R, Tran T, et al. Protein kinase G and focal adhesion kinase converge on Src/Akt/β-catenin signaling module in osteoblast mechanotransduction. J Biol Chem. 2012;287:21509–19.[PMC free article] [PubMed]
51. Marathe N, Rangaswami H, Zhuang S, et al. Pro-survival effects of 17β-estradiol on osteocytes are mediated by nitric oxide/cGMP via differential actions of cGMP-dependent protein kinases I and II. J Biol Chem. 2012;287:978–88. [PMC free article] [PubMed]
52. Rangaswami H, Schwappacher R, Marathe N, et al. Cyclic GMP and protein kinase G control a Src-containing mechanosome in osteoblasts. Sci Signal. 2010;3:ra91. [PMC free article] [PubMed]
53. Rangaswami H, Marathe N, Zhuang S, et al. Type II cGMP-dependent protein kinase mediates osteoblast mechanotransduction. J Biol Chem. 2009;284:14796–808. [PMC free article] [PubMed]
54. Saura M, Tarin C, Zaragoza C. Recent insights into the implication of nitric oxide in osteoblast differentiation and proliferation during bone development. ScientificWorldJournal. 2010;10:624–32.[PubMed]
55. Zaragoza C, López-Rivera E, García-Rama C, et al. Cbfa-1 mediates nitric oxide regulation of MMP-13 in osteoblasts. J Cell Sci. 2006;119:1896–902. [PubMed]
56. Wang DH, Hu YS, Du JJ, et al. Ghrelin stimulates proliferation of human osteoblastic TE85 cells viaNO/cGMP signaling pathway. Endocrine. 2009;35:112–7. [PubMed]
57. Simkó M. Cell type specific redox status is responsible for diverse electromagnetic field effects. Curr Med Chem. 2007;14:1141–52. [PubMed]
58. Consales C, Merla C, Marino C, et al. Electromagnetic fields, oxidative stress, and neurodegeneration. Int J Cell Biol. 2012;2012:683897. [PMC free article] [PubMed]
59. Johansson O. Disturbance of the immune system by electromagnetic fields-A potentially underlying cause for cellular damage and tissue repair reduction which could lead to disease and impairment.Pathophysiology. 2009;16:157–77. [PubMed]
60. Kovacic P, Somanathan R. Electromagnetic fields: mechanism, cell signaling, other bioprocesses, toxicity, radicals, antioxidants and beneficial effects. J Recept Signal Transduct Res. 2010;30:214–26.[PubMed]
61. Wolf FI, Torsello A, Tedesco B, et al. 50-Hz extremely low frequency electromagnetic fields enhance cell proliferation and DNA damage: possible involvement of a redox mechanism. Biochim Biophys Acta.2005;1743:120–9. [PubMed]
62. Iakimenko IL, Sidorik EP, Tsybulin AS. Metabolic changes in cells under electromagnetic radiation of mobile communication systems. Ukr Biokhim Zh. 2011;83:20–8. [PubMed]
63. Jing J, Yuhua Z, Xiao-qian Y, et al. The influence of microwave radiation from cellular phone on fetal rat brain. Electromagn Biol Med. 2012;31:57–66. [PubMed]
64. Esmekaya MA, Ozer C, Seyhan N. 900 MHz pulse-modulated radiofrequency radiation induces oxidative stress on heart, lung, testis and liver tissues. Gen Physiol Biophys. 2011;30:84–9. [PubMed]
65. Aydin B, Akar A. Effects of a 900-MHz electromagnetic field on oxidative stress parameters in rat lymphoid organs, polymorphonuclear leukocytes and plasma. Arch Med Res. 2011;42:261–7. [PubMed]
66. Guler G, Turkozer Z, Tomruk A, et al. The protective effects of N-acetyl-L-cysteine and epigallocatechin-3-gallate on electric field-induced hepatic oxidative stress. Int J Radiat Biol. 2008;84:669–80. [PubMed]
67. Guney M, Ozguner F, Oral B, et al. 900 MHz radiofrequency-induced histopathologic changes and oxidative stress in rat endometrium: protection by vitamins E and C. Toxicol Ind Health. 2007;23:411–20.[PubMed]
68. Sypniewska RK, Millenbaugh NJ, Kiel JL, et al. Protein changes in macrophages induced by plasma from rats exposed to 35 GHz millimeter waves. Bioelectromagnetics. 2010;31:656–63. [PubMed]
69. Grigoriev YG, Mikhailov VF, Ivanov AA, et al. Autoimmune processes after long-term low-level exposure to electromagnetic fields part 4. Oxidative intracellular stress response to the long-term rat exposure to nonthermal RF EMF. Biophysics. 2010;55:1054–8.
70. Erdal N, Gürgül S, Tamer L, et al. Effects of long-term exposure of extremely low frequency magnetic field on oxidative/nitrosative stress in rat liver. J Radiat Res. 2008;49:181–7. [PubMed]
71. Ahuja YR, Vijayashree B, Saran R, et al. In vitro effects of low-level, low-frequency electromagnetic fields on DNA damage in human leucocytes by comet assay. Indian J Biochem Biophys. 1999;36:318–22.[PubMed]
72. Amara S, Douki T, Ravanat JL, et al. Influence of a static magnetic field (250 mT) on the antioxidant response and DNA integrity in THP1 cells. Phys Med Biol. 2007;52:889–98. [PubMed]
73. Focke F, Schuermann D, Kuster N, et al. DNA fragmentation in human fibroblasts under extremely low frequency electromagnetic field exposure. Mutat Res. 2010;683:74–83. [PubMed]
74. Franzellitti S, Valbonesi P, Ciancaglini N, et al. Transient DNA damage induced by high-frequency electromagnetic fields (GSM 1.8 GHz) in the human trophoblast HTR-8/SVneo cell line evaluated with the alkaline comet assay. Mutat Res. 2010;683:35–42. [PubMed]
75. Garaj-Vrhovac V, Gajski G, Pažanin S, et al. Assessment of cytogenetic damage and oxidative stress in personnel occupationally exposed to the pulsed microwave radiation of marine radar equipment. Int J Hyg Environ Health. 2011;214:59–65. [PubMed]
76. Hong R, Zhang Y, Liu Y, et al. Effects of extremely low frequency electromagnetic fields on DNA of testicular cells and sperm chromatin structure in mice. Zhonghua Lao Dong Wei Sheng Zhi Ye Bing Za Zhi.2005;23:414–7. [Article in Chinese] [PubMed]
77. Ivancsits S, Diem E, Pilger A, et al. Induction of DNA strand breaks by intermittent exposure to extremely-low-frequency electromagnetic fields in human diploid fibroblasts. Mutat Res. 2002;519:1–13.[PubMed]
78. Ivancsits S, Diem E, Jahn O, et al. Intermittent extremely low frequency electromagnetic fields cause DNA damage in a dose-dependent way. Int Arch Occup Environ Health. 2003;76:431–6. [PubMed]
79. Ivancsits S, Pilger A, Diem E, et al. Cell type-specific genotoxic effects of intermittent extremely low-frequency electromagnetic fields. Mutat Res. 2005;583:184–8. [PubMed]
80. Kesari KK, Behari J, Kumar S. Mutagenic response of 2.45 GHz radiation exposure on rat brain. Int J Radiat Biol. 2010;86:334–43. [PubMed]
81. Lai H, Singh NP. Melatonin and a spin-trap compound block radiofrequency electromagnetic radiation-induced DNA strand breaks in rat brain cells. Bioelectromagnetics. 1997;18:446–54. [PubMed]
82. Lai H, Singh NP. Magnetic-field-induced DNA strand breaks in brain cells of the rat. Environ Health Perspect. 2004;112:687–94. [PMC free article] [PubMed]
83. Lee JW, Kim MS, Kim YJ, et al. Genotoxic effects of 3 T magnetic resonance imaging in cultured human lymphocytes. Bioelectromagnetics. 2011;32:535–42. [PubMed]
84. Paulraj R, Behari J. Single strand DNA breaks in rat brain cells exposed to microwave radiation. Mutat Res. 2006;596:76–80. [PubMed]
85. Romeo S, Zeni L, Sarti M, et al. DNA electrophoretic migration patterns change after exposure of Jurkat cells to a single intense nanosecond electric pulse. PLoS ONE. 2011;6:e28419. [PMC free article] [PubMed]
86. Schwarz C, Kratochvil E, Pilger A, et al. Radiofrequency electromagnetic fields (UMTS, 1,950 MHz) induce genotoxic effects in vitro in human fibroblasts but not in lymphocytes. Int Arch Occup Environ Health. 2008;81:755–67. [PubMed]
87. Svedenstål BM, Johanson KJ, Mattsson MO, et al. DNA damage, cell kinetics and ODC activities studied in CBA mice exposed to electromagnetic fields generated by transmission lines. In Vivo.1999;13:507–13. [PubMed]
88. Svedenstål BM, Johanson KJ, Mild KH. DNA damage induced in brain cells of CBA mice exposed to magnetic fields. In Vivo. 1999;13:551–2. [PubMed]
89. Trosić I, Pavicić I, Milković-Kraus S, et al. Effect of electromagnetic radiofrequency radiation on the rats’ brain, liver and kidney cells measured by comet assay. Coll Antropol. 2011;35:1259–64. [PubMed]
90. Burdak-Rothkamm S, Rothkamm K, Folkard M, et al. DNA and chromosomal damage in response to intermittent extremely low-frequency magnetic fields. Mutat Res. 2009;672:82–9. [PubMed]
91. Fairbairn DW, O'Neill KL. The effect of electromagnetic field exposure on the formation of DNA single strand breaks in human cells. Cell Mol Biol (Noisy-le-grand) 1994;40:561–7. [PubMed]
92. Fiorani M, Cantoni O, Sestili P, et al. Electric and/or magnetic field effects on DNA structure and function in cultured human cells. Mutat Res. 1992;282:25–9. [PubMed]
93. Malyapa RS, Ahern EW, Straube WL, et al. Measurement of DNA damage after exposure to 2450 MHz electromagnetic radiation. Radiat Res. 1997;148:608–17. [PubMed]
94. McNamee JP, Bellier PV, Chauhan V, et al. Evaluating DNA damage in rodent brain after acute 60 Hz magnetic-field exposure. Radiat Res. 2005;164:791–7. [PubMed]
95. Scarfí MR, Sannino A, Perrotta A, et al. Evaluation of genotoxic effects in human fibroblasts after intermittent exposure to 50 Hz electromagnetic fields: a confirmatory study. Radiat Res. 2005;164:270–6.[PubMed]
96. Stronati L, Testa A, Villani P, et al. Absence of genotoxicity in human blood cells exposed to 50 Hz magnetic fields as assessed by comet assay, chromosome aberration, micronucleus, and sister chromatid exchange analyses. Bioelectromagnetics. 2004;25:41–8. [PubMed]
97. Testa A, Cordelli E, Stronati L, et al. Evaluation of genotoxic effect of low level 50 Hz magnetic fields on human blood cells using different cytogenetic assays. Bioelectromagnetics. 2004;25:613–9. [PubMed]
98. Szabó G, Bährle S. Role of nitrosative stress and poly(ADP-ribose) polymerase activation in myocardial reperfusion injury. Curr Vasc Pharmacol. 2005;3:215–20. [PubMed]
99. Moon HK, Yang ES, Park JW. Protection of peroxynitrite-induced DNA damage by dietary antioxidants. Arch Pharm Res. 2006;29:213–7. [PubMed]
100. Sakihama Y, Maeda M, Hashimoto M, et al. Beetroot betalain inhibits peroxynitrite-mediated tyrosine nitration and DNA strand damage. Free Radic Res. 2012;46:93–9. [PubMed]
101. Hybertson BM, Gao B, Bose SK, et al. Oxidative stress in health and disease: the therapeutic potential of Nrf2 activation. Mol Aspects Med. 2011;32:234–46. [PubMed]
102. Ridley AJ, Paterson HF, Johnston CL, et al. The small GTP-binding protein rac regulates growth factor-induced membrane ruffling. Cell. 1992;70:401–10. [PubMed]
103. Wennström S, Hawkins P, Cooke F, et al. Activation of phosphoinositide 3-kinase is required for PDGF-stimulated membrane ruffling. Curr Biol. 1994;4:385–93. [PubMed]
104. Joucla S, Yvert B. Modeling of extracellular neural stimulation: from basic understanding to MEA-based applications. J Physiol Paris. 2012;106:146–58. [PubMed]
105. Pashut T, Wolfus S, Friedman A, et al. Mechanisms of magnetic stimulation of central nervous system neurons. PLoS Comput Biol. 2011;7:e1002022. [PMC free article] [PubMed]
106. Fatemi-Ardekani A. Transcranial magnetic stimulation: physics, electrophysiology, and applications.Crit Rev Biomed Eng. 2008;36:375–412. [PubMed]
107. Silva S, Basser PJ, Miranda PC. Elucidating the mechanisms and loci of neuronal excitation by transcranial magnetic stimulation using a finite element model of a cortical sulcus. Clin Neurophysiol.2008;119:2405–13. [PMC free article] [PubMed]
108. Radman T, Ramos RL, Brumberg JC, et al. Role of cortical cell type and morphology in subthreshold and suprathreshold uniform electric field stimulation in vitro. Brain Stimul. 2009;2:215–28.[PMC free article] [PubMed]
109. Minelli TA, Balduzzo M, Milone FF, et al. Modeling cell dynamics under mobile phone radiation. Nonlinear cell dynamics under mobile phone radiation. Nonlinear Dynamics Psychol Life Sci. 2007;11:197–218. [PubMed]
110. Saunders RD, Jefferys JGR. A neurobiological basis for ELF guidelines. Health Phys. 2007;92:596–603. [PubMed]
111. Havas M. Dirty electricity elevates blood sugar among electrically sensitive diabetics and may explain brittle diabetes. Electromagn Biol Med. 2008;27:135–46. [PMC free article] [PubMed]
112. Havas M. Electromagnetic hypersensitivity: biological effects of dirty electricity with emphasis on diabetes and multiple sclerosis. Electromagn Biol Med. 2006;25:259–68. [PubMed]
113. de Vochta F. “Dirty electricity”: what, where, and should we care? J Expo Sci Environ Epidemiol.2010;20:399–405. [PubMed]
114. Friedman J, Kraus S, Hauptman Y, et al. Mechanism of short-term ERK activation by electromagnetic fields at mobile phone frequencies. Biochem J. 2007;405:559–68. [PMC free article] [PubMed]
115. Desai NR, Kesari KK, Agarwal A. Pathophysiology of cell phone radiation: oxidative stress and carcinogenesis with focus on the male reproductive system. Reproduct Biol Endocrinol. 2009;7:114. doi:10.1186/1477-7827-7-114. [PMC free article] [PubMed]
116. Wyatt CN, Weir EK, Peers C. Diphenylamine iodonium blocks K+ and Ca2+ currents in type I cells isolated from the rat carotid body. Neurosci Lett. 1994;172:63–6. [PubMed]
Articles from Journal of Cellular and Molecular Medicine are provided here courtesy of Blackwell Publishing
EMF responses blocked or lowered by calcium channel blockers
In a recent study, Pilla [27] showed that an increase in intracellular Ca2+ must have occurred almost immediately after EMF exposure, producing a Ca2+/calmodulin-dependent increase in nitric oxide occurring in less than 5 sec. Although Pilla [27] did not test whether VGCC stimulation was involved in his study, there are few alternatives that can produce such a rapid Ca2+ response, none of which has been implicated in EMF responses. Other studies, each involving VGCCs, summarized in Table 1, also showed rapid Ca2+increases following EMF exposure [8, 16, 17, 19, 21]. The rapidity of these responses rule out many types of regulatory interactions as being involved in producing the increased VGCC activity following EMF exposure and suggests, therefore, that VGCC stimulation in the plasma membrane is directly produced by EMF exposure.
Go to:
Possible modes of action following VGCC stimulation
The increased intracellular Ca2+ produced by such VGCC activation may lead to multiple regulatory responses, including the increased nitric oxide levels produced through the action of the two Ca2+/calmodulin-dependent nitric oxide synthases, nNOS and eNOS. Increased nitric oxide levels typically act in a physiological context through increased synthesis of cGMP and subsequent activation of protein kinase G [28, 29]. In contrast, in most pathophysiological contexts, nitric oxide reacts with superoxide to form peroxynitrite, a potent non-radical oxidant [30, 31], which can produce radical products, including hydroxyl radical and NO2 radical [32].
Go to:
Therapeutic bone-growth stimulation via Ca2+/nitric oxide/cGMP/protein kinase G
An example of a therapeutic effect for bone repair of EMF exposure in various medical situations includes increasing osteoblast differentiation and maturation and has been reviewed repeatedly [33–44]. The effects of EMF exposure on bone cannot be challenged, although there is still considerable question about the best ways to apply this clinically [33–44]. Our focus, here, is to consider possible mechanisms of action. Multiple studies have implicated increased Ca2+ and nitric oxide in the EMF stimulation of bone growth [44–49]; three have also implicated increased cGMP and protein kinase G activity [46, 48, 49]. In addition, studies on other regulatory stimuli leading to increased bone growth have also implicated increased cGMP levels and protein kinase G in this response [50–56]. In summary, then, it can be seen from the above that there is a very well-documented action of EMFs in stimulating osteoblasts and bone growth. The available data, although limited, support the action of the main pathway involved in physiological responses to Ca2+ and nitric oxide, namely Ca2+/nitric oxide/cGMP/protein kinase G in producing such stimulation.
Go to:
Ca2+/nitric oxide/peroxynitrite and pathophysiological responses to EMF exposures: the example of single-strand DNA breaks
As was noted above, most of the pathophysiological effects of nitric oxide are mediated through peroxynitrite elevation and consequent oxidative stress. There are many reviews and other studies, implicating oxidative stress in generating pathophysiological effects of EMF exposure [see for example [57–64]]. In some of these studies, the rise in oxidative stress markers parallels the rise in nitric oxide, suggesting a peroxynitrite-mediated mechanism [64–67].
Peroxynitrite elevation is usually measured through a marker of peroxynitrite-mediated protein nitration, 3-nitrotyrosine (3-NT). There are four studies where 3-NT levels were measured before and after EMF exposure [66, 68–70]. Each of these studies provides some evidence supporting the view that EMF exposure increases levels of peroxynitrite and therefore 3-NT levels [66, 68–70]. Although these cannot be taken as definitive, when considered along with the evidence on oxidative stress and elevated nitric oxide production in response to EMF exposure, they strongly suggest a peroxynitrite-mediated mechanism of oxidative stress in response to EMFs.
Such a peroxynitrite-mediated mechanism may explain the many studies showing the single-stranded breaks in DNA, as shown by alkaline comet assays or the similar microgel electrophoresis assay, following EMF exposures in most such studies [71–89], but not in all [90–97]. Some of the factors that are reported to influence whether such DNA single-strand breaks are detected after EMF exposure include the type of cell studied [79, 86], dosage of EMF exposure [78] and the type of EMF exposure studied [73, 77]. Oxidative stress and free radicals have roles, both because there is a concomitant increase in oxidative stress and because antioxidants have been shown to greatly lower the generation of DNA single-strand breaks following EMF exposure [72, 75, 81, 82] as has also been shown for peroxynitrite-mediated DNA breaks produced under other conditions. It has also been shown that one can block the generation of DNA single-strand breaks with a nitric oxide synthase inhibitors [82].
Peroxynitrite has been shown to produce single-strand DNA breaks [98–100], a process that is inhibited by many but not all antioxidants [99, 100]. It can be seen from this that the data on generation of single-strand DNA breaks, although quite limited, support a mechanism involving nitric oxide/peroxynitrite/free radical (oxidative stress). Although the data on the possible role of peroxynitrite in EMF-induced DNA single-strand breaks are limited, what data are available supports such a peroxynitrite role.
Go to:
Discussion and conclusions
How do EMFs composed of low-energy photons produce non-thermal biological changes, both pathophysiological and, in some cases, potentially therapeutic, in humans and higher animals? It may be surprising that the answer to this question has been hiding in plain sight in the scientific literature. However, in this era of highly focused and highly specialized science, few of us have the time to read the relevant literature, let alone organize the information found within it in useful and critical ways.
This study shows that:
- Twenty-three different studies have found that such EMF exposures act via activation of VGCCs, such that VGCC channel blockers can prevent responses to such exposures (Table 1). Most of the studies implicate L-type VGCCs in these responses, but there are also other studies implicating three other classes of VGCCs.
- Both extremely low frequency fields, including 50/60 cycle exposures, and microwave EMF range exposures act via activation of VGCCs. So do static electric fields, static magnetic fields and nanosecond pulses.
- Voltage-gated calcium channel stimulation leads to increased intracellular Ca2+, which can act in turn to stimulate the two calcium/calmodulin-dependent nitric oxide synthases and increase nitric oxide. It is suggested here that nitric oxide may act in therapeutic/potentially therapeutic EMF responses via its main physiological pathway, stimulating cGMP and protein kinase G. It is also suggested that nitric oxide may act in pathophysiological responses to EMF exposure, by acting as a precursor of peroxynitrite, producing both oxidative stress and free radical breakdown products.
- The interpretation in three above is supported by two specific well-documented examples of EMF effects. Electromagnetic fields stimulation of bone growth, modulated through EMF stimulation of osteoblasts, appears to involve an elevation/nitric oxide/protein kinase G pathway. In contrast to that, it seems likely that the EMF induction of single-stranded DNA breaks involves a Ca2+/elevation/nitric oxide/peroxynitrite/free radical (oxidative stress) pathway.
The possible role of peroxynitrite as opposed to protein kinase G in producing pathophysiological responses to EMF exposure raises the question of whether there are practical approaches to avoiding such responses? Typically peroxynitrite levels can be highly elevated when both of its precursors, nitric oxide and superoxide, are high. Consequently, agents that lower nitric oxide synthase activity and agents that raise superoxide dismutases (SODs, the enzymes that degrade superoxide) such as phenolics and other Nrf2 activators that induce SOD activity [101], as well as calcium channel blockers may be useful. Having said that, this is a complex area, where other approaches should be considered, as well.
Although the various EMF exposures as well as static electrical field exposures can act to change the electrical voltage-gradient across the plasma membrane and may, therefore, be expected to stimulate VGCCs through their voltage-gated properties, it may be surprising that static magnetic fields also act to activate VGCCs because static magnetic fields do not induce electrical changes on static objects. However, cells are far from static. Such phenomena as cell ruffling [102],[103] may be relevant, where thin cytoplasmic sheets bounded on both sides by plasma membrane move rapidly. Such rapid movement of the electrically conducting cytoplasm, may be expected to influence the electrical charge across the plasma membrane, thus potentially stimulating the VGCCs.
Earlier modelling of electrical effects across plasma membranes of EMF exposures suggested that such electrical effects were likely to be too small to explain EMF effects at levels reported to produce biological changes (see, for example [22]). However, more recent and presumably more biologically plausible modelling have suggested that such electrical effects may be much more substantial [104–109] and may, therefore, act to directly stimulate VGCCs.
Direct stimulation of VGCCs by partial depolarization across the plasma membrane is suggested by the following observations discussed in this review:
- The very rapid, almost instantaneous increase in intracellular Ca2+ found in some studies following EMF exposure [8, 16, 17, 19, 21, 27]. The rapidity here means that most, if not all indirect, regulatory effects can be ruled out.
- The fact that not just L-type, but three additional classes of VGCCs are implicated in generating biological responses to EMF exposure (Table 1), suggesting that their voltage-gated properties may be a key feature in their ability to respond to EMFs.
- Most, if not all, EMF effects are blocked by VGCC channel blockers (Table 1).
- Modelling of EMF effects on living cells suggests that plasma membrane voltage changes may have key roles in such effects [104–109]. Saunders and Jefferys stated [110] that ‘It is well established that electric fields … or exposure to low frequency magnetic fields, will, if of sufficient magnitude, excite nerve tissue through their interactions with … voltage gated ion channels’. They further state [110] that this is achieved by direct effects on the electric dipole voltage sensor within the ion channel.
The only detailed alternative to the mechanism of non-thermal EMF effects discussed here, to my knowledge, is the hypothesis of Friedman et al. [114] and supported by Desai et al. [115] where the apparent initial response to EMF exposure was proposed to be NADH oxidase activation, leading to oxidative stress and downstream regulatory effects. Although they provide some correlative evidence for a possible role of NADH oxidase [114], the only causal evidence is based on a presumed specific inhibitor of NADH oxidase, diphenyleneiodonium (DPI). However, DPI has been shown to be a non-specific cation channel blocker [116], clearly showing a lack of such specificity and suggesting that it may act, in part, as a VGCC blocker. Consequently, a causal role for NADH oxidase in responses to EMF exposure must be considered to be undocumented.
In summary, the non-thermal actions of EMFs composed of low-energy photons have been a great puzzle, because such photons are insufficiently energetic to directly influence the chemistry of cells. The current review provides support for a pathway of the biological action of ultralow frequency and microwave EMFs, nanosecond pulses and static electrical or magnetic fields: EMF activation of VGCCs leads to rapid elevation of intracellular Ca2+, nitric oxide and in some cases at least, peroxynitrite. Potentially therapeutic effects may be mediated through the Ca2+/nitric oxide/cGMP/protein kinase G pathway. Pathophysiological effects may be mediated through the Ca2+/nitric oxide/peroxynitrite pathway. Other Ca2+-mediated effects may have roles as well, as suggested by Xu et al. [26].
Go to:
Conflicts of interest
The author confirms that there are no conflicts of interest.
Go to:
References
1. Walleczek J. Electromagnetic field effects on cells of the immune system: the role of calcium signaling.FASEB J. 1992;6:3177–85. [PubMed]
2. Cadossi R, Emilia G, Ceccherelli G. 1988 Lymphocytes and pulsing magnetic fields. In: Marino EE, et al., editors. Modern bioelectricity. New York: Dekker; 1998. pp. 451–96.
3. Papatheofanis FJ. Use of calcium channel antagonists as magnetoprotective agents. Radiat Res.1990;122:24–8. [PubMed]
4. Catterall WA. Structure and regulation of voltage-gated Ca2+ channels. Annu Rev Cell Dev Biol.2000;16:521–55. [PubMed]
5. Morgado-Valle C, Verdugo-Díaz L, García DE, et al. The role of voltage-gated Ca2+ channels in neurite growth of cultured chromaffin cells induced by extremely low frequency (ELF) magnetic field stimulation.Cell Tissue Res. 1998;291:217–30. [PubMed]
6. Lorich DG, Brighton CT, Gupta R, et al. Biochemical pathway mediating the response of bone cells to capacitive coupling. Clin Orthop Relat Res. 1998:246–56. [PubMed]
7. Gobba F, Malagoli D, Ottaviani E. Effects of 50 Hz magnetic fields on fMLP-induced shape changes in invertebrate immunocytes: the role of calcium ion channels. Bioelectromagnetics. 2003;24:277–82.[PubMed]
8. Lisi A, Ledda M, Rosola E, et al. Extremely low frequency electromagnetic field exposure promotes differentiation of pituitary corticotrope-derived AtT20 D16V cells. Bioelectromagnetics. 2006;27:641–51.[PubMed]
9. Piacentini R, Ripoli C, Mezzogori D, et al. Extremely low-frequency electromagnetic fields promote in vitro neurogenesis via upregulation of Ca(v)1-channel activity. J Cell Physiol. 2008;215:129–39. [PubMed]
10. Morris CE, Skalak TC. Acute exposure to a moderate strength static magnetic field reduces edema formation in rats. Am J Physiol Heart Circ Physiol. 2008;294:H50–7. [PubMed]
11. Ghibelli L, Cerella C, Cordisco S, et al. NMR exposure sensitizes tumor cells to apoptosis. Apoptosis.2006;11:359–65. [PubMed]
12. Fanelli C, Coppola S, Barone R, et al. Magnetic fields increase cell survival by inhibiting apoptosis viamodulation of Ca2+ influx. FASEB J. 1999;13:95–102. [PubMed]
13. Jeong JH, Kum C, Choi HJ, et al. Extremely low frequency magnetic field induces hyperalgesia in mice modulated by nitric oxide synthesis. Life Sci. 2006;78:1407–12. [PubMed]
14. Vernier PT, Sun Y, Chen MT, et al. Nanosecond electric pulse-induced calcium entry into chromaffin cells. Bioelectrochemistry. 2008;73:1–4. [PubMed]
15. Kim IS, Song JK, Song YM, et al. Novel effect of biphasic electric current on in vitro osteogenesis and cytokine production in human mesenchymal stromal cells. Tissue Eng Part A. 2009;15:2411–22. [PubMed]
16. Höjevik P, Sandblom J, Galt S, et al. Ca2+ ion transport through patch-clamped cells exposed to magnetic fields. Bioelectromagnetics. 1995;16:33–40. [PubMed]
17. Barbier E, Vetret B, Dufy B. Stimulation of Ca2+ influx in rat pituitary cells under exposure to a 50 Hz magnetic field. Bioelectromagnetics. 1996;17:303–11. [PubMed]
18. Grassi C, D'Ascenzo M, Torsello A, et al. Effects of 50 Hz electromagnetic fields on voltage-gated Ca2+channels and their role in modulation of neuroendocrine cell proliferation and death. Cell Calcium.2004;35:307–15. [PubMed]
19. Craviso GL, Choe S, Chatterjee P, et al. Nanosecond electric pulses: a novel stimulus for triggering Ca2+influx into chromaffin cells via voltage-gated Ca2+ channels. Cell Mol Neurobiol. 2010;30:1259–65.[PubMed]
20. Marchionni I, Paffi A, Pellegrino M, et al. Comparison between low-level 50 Hz and 900 MHz electromagnetic stimulation on single channel ionic currents and on firing frequency in dorsal root ganglion isolated neurons. Biochim Biophys Acta. 2006;1758:597–605. [PubMed]
21. Rao VS, Titushkin IA, Moros EG, et al. Nonthermal effects of radiofrequency-field exposure on calcium dynamics in stem cell-derived neuronal cells: elucidation of calcium pathways. Radiat Res. 2008;169:319–29. [PubMed]
22. Adair RK, Astumian RD, Weaver JC. Detection of weak electric fields by sharks, rays and skates.Chaos. 1998;8:576–87. [PubMed]
23. Constable PA. Nifedipine alters the light-rise of the electro-oculogram in man. Fraefes Arch Clin Exp Ophthalmol. 2011;249:677–84. [PubMed]
24. Gmitrov J, Ohkuba C. Verapamil protective effect on natural and artificial magnetic field cardiovascular impact. Bioelectromagnetics. 2002;23:531–41. [PubMed]
25. Kindzelskii AL, Petty HR. Ion channel clustering enhances weak electric field detection by neutrophils: apparent role of SKF96365-sensitive cation channels and myeloperoxidase trafficking cellular responses. Eur Biophys J. 2005;35:1–26. [PubMed]
26. Xu J, Wang W, Clark CC, et al. Signal transduction in electrically stimulated articular chondrocytes involves translocation of extracellular calcium through voltage-gated channels. Osteoarthritis Cartilage.2009;17:397–405. [PubMed]
27. Pilla AA. Electromagnetic fields instantaneously modulate nitric oxide signaling in challenged biological systems. Biochem Biophys Res Commun. 2012;426:330–3. [PubMed]
28. McDonald LJ, Murad F. Nitric oxide and cyclic GMP signaling. Proc Soc Exp Biol Med. 1996;211:1–6.[PubMed]
29. Francis SH, Busch JL, Corbin JD, et al. cGMP-dependent protein kinases and cGMP phosphodiesterases in nitric oxide and cGMP action. Pharmacol Rev. 2010;62:525–63. [PMC free article] [PubMed]
30. Pacher P, Beckman JS, Liaudet L. Nitric oxide and peroxynitrite in health and disease. Physiol Rev.2007;87:315–424. [PMC free article] [PubMed]
31. Pryor WA, Squadrito GL. The chemistry of peroxynitrite: a product from the reaction of nitric oxide with superoxide. Am J Physiol. 1995;268:L699–722. [PubMed]
32. Lymar SV, Khairutdinov RF, Hurst JK. Hydroxyl radical formation by O-O bond homolysis in peroxynitrous acid. Inorg Chem. 2003;42:5259–66. [PubMed]
33. Ryabi JT. Clinical effects of electromagnetic fields on fracture healing. Clin Orthop Relat Res.1998;355(Suppl. l):S205–15. [PubMed]
34. Oishi M, Onesti ST. Electrical bone graft stimulation for spinal fusion: a review. Neurosurgery.2000;47:1041–55. [PubMed]
35. Aaron RK, Ciombor DM, Simon BJ. Treatment of nonunions with electric and electromagnetic fields.Clin Orthop Relat Res. 2004;10:579–93. [PubMed]
36. Goldstein C, Sprague S, Petrisor BA. Electrical stimulation for fracture healing: current evidence. J Orthop Trauma. 2010;24(Suppl. 1):S62–5. [PubMed]
37. Demitriou R, Babis GC. Biomaterial osseointegration enhancement with biophysical stimulation. J Musculoskelet Neuronal Interact. 2007;7:253–65. [PubMed]
38. Griffin XL, Warner F, Costa M. The role of electromagnetic stimulation in the management of established non-union lf long bone fractures: what is the evidence? Injury. 2008;39:419–29. [PubMed]
39. Huang LQ, He HC, He CQ, et al. Clinical update of pulsed electromagnetic fields on osteroporosis. Chin Med J. 2008;121:2095–9. [PubMed]
40. Groah SL, Lichy AM, Libin AV, et al. Intensive electrical stimulation attenuates femoral bone loss in acute spinal cord injury. PM R. 2010;2:1080–7. [PubMed]
41. Schidt-Rohlfing B, Silny J, Gavenis K, et al. Electromagnetic fields, electric current and bone healing – what is the evidence? Z Orthop Unfall. 2011;149:265–70. [PubMed]
42. Griffin XL, Costa ML, Parsons N, et al. Electromagnetic field stimulation for treating delayed union or non-union of long bone fractures in adults. Cochrane Database Syst Rev. 2011:CDO08471. doi:10.1002/14651858.CD008471.pub2. [PubMed]
43. Chalidis B, Sachinis N, Assiotis A, et al. Stimulation of bone formation and fracture healing with pulsed electromagnetic fields: biologic responses and clinical implications. Int J Immunopathol Pharmacol.2011;24(1 Suppl. 2):17020. [PubMed]
44. Zhong C, Zhao TF, Xu ZJ, et al. Effects of electromagnetic fields on bone regeneration in experimental and clinical studies: a review of the literature. Chin Med J. 2012;125:367–72. [PubMed]
45. Diniz P, Soejima K, Ito G. Nitric oxide mediates the effects of pulsed electromagnetic field stimulation on the osteoblast proliferation and differentiation. Nitric Oxide. 2002;7:18–23. [PubMed]
46. Fitzsimmons RJ, Gordon SL, Ganey T, et al. A pulsing electric field (PEF) increases human chondrocyte proliferation through a transduction pathway involving nitric oxide signaling. J Orthopaedic Res.2008;26:854–9. [PubMed]
47. Lin H-Y, Lin Y-J. In vitro effects of low frequency electromagnetic fields on osteoblast proliferation and maturation in an inflammatory environment. Bioelectromagnetics. 2011;32:552–60. [PubMed]
48. Cheng G, Zhai Y, Chen K, et al. Sinusoidal electromagnetic field stimulates rat osteoblast differentiation and maturation via activation of NO-cGMP-PKG pathway. Nitric Oxide. 2011;25:316–25. [PubMed]
49. Pilla A, Fitzsimmons R, Muehsam D, et al. Electromagnetic fields as first messenger in biological signaling: application to calmodulin-dependent signaling in tissue repair. Biochim Biophys Acta.2011;1810:1236–45. [PubMed]
50. Rangaswami H, Schwappacher R, Tran T, et al. Protein kinase G and focal adhesion kinase converge on Src/Akt/β-catenin signaling module in osteoblast mechanotransduction. J Biol Chem. 2012;287:21509–19.[PMC free article] [PubMed]
51. Marathe N, Rangaswami H, Zhuang S, et al. Pro-survival effects of 17β-estradiol on osteocytes are mediated by nitric oxide/cGMP via differential actions of cGMP-dependent protein kinases I and II. J Biol Chem. 2012;287:978–88. [PMC free article] [PubMed]
52. Rangaswami H, Schwappacher R, Marathe N, et al. Cyclic GMP and protein kinase G control a Src-containing mechanosome in osteoblasts. Sci Signal. 2010;3:ra91. [PMC free article] [PubMed]
53. Rangaswami H, Marathe N, Zhuang S, et al. Type II cGMP-dependent protein kinase mediates osteoblast mechanotransduction. J Biol Chem. 2009;284:14796–808. [PMC free article] [PubMed]
54. Saura M, Tarin C, Zaragoza C. Recent insights into the implication of nitric oxide in osteoblast differentiation and proliferation during bone development. ScientificWorldJournal. 2010;10:624–32.[PubMed]
55. Zaragoza C, López-Rivera E, García-Rama C, et al. Cbfa-1 mediates nitric oxide regulation of MMP-13 in osteoblasts. J Cell Sci. 2006;119:1896–902. [PubMed]
56. Wang DH, Hu YS, Du JJ, et al. Ghrelin stimulates proliferation of human osteoblastic TE85 cells viaNO/cGMP signaling pathway. Endocrine. 2009;35:112–7. [PubMed]
57. Simkó M. Cell type specific redox status is responsible for diverse electromagnetic field effects. Curr Med Chem. 2007;14:1141–52. [PubMed]
58. Consales C, Merla C, Marino C, et al. Electromagnetic fields, oxidative stress, and neurodegeneration. Int J Cell Biol. 2012;2012:683897. [PMC free article] [PubMed]
59. Johansson O. Disturbance of the immune system by electromagnetic fields-A potentially underlying cause for cellular damage and tissue repair reduction which could lead to disease and impairment.Pathophysiology. 2009;16:157–77. [PubMed]
60. Kovacic P, Somanathan R. Electromagnetic fields: mechanism, cell signaling, other bioprocesses, toxicity, radicals, antioxidants and beneficial effects. J Recept Signal Transduct Res. 2010;30:214–26.[PubMed]
61. Wolf FI, Torsello A, Tedesco B, et al. 50-Hz extremely low frequency electromagnetic fields enhance cell proliferation and DNA damage: possible involvement of a redox mechanism. Biochim Biophys Acta.2005;1743:120–9. [PubMed]
62. Iakimenko IL, Sidorik EP, Tsybulin AS. Metabolic changes in cells under electromagnetic radiation of mobile communication systems. Ukr Biokhim Zh. 2011;83:20–8. [PubMed]
63. Jing J, Yuhua Z, Xiao-qian Y, et al. The influence of microwave radiation from cellular phone on fetal rat brain. Electromagn Biol Med. 2012;31:57–66. [PubMed]
64. Esmekaya MA, Ozer C, Seyhan N. 900 MHz pulse-modulated radiofrequency radiation induces oxidative stress on heart, lung, testis and liver tissues. Gen Physiol Biophys. 2011;30:84–9. [PubMed]
65. Aydin B, Akar A. Effects of a 900-MHz electromagnetic field on oxidative stress parameters in rat lymphoid organs, polymorphonuclear leukocytes and plasma. Arch Med Res. 2011;42:261–7. [PubMed]
66. Guler G, Turkozer Z, Tomruk A, et al. The protective effects of N-acetyl-L-cysteine and epigallocatechin-3-gallate on electric field-induced hepatic oxidative stress. Int J Radiat Biol. 2008;84:669–80. [PubMed]
67. Guney M, Ozguner F, Oral B, et al. 900 MHz radiofrequency-induced histopathologic changes and oxidative stress in rat endometrium: protection by vitamins E and C. Toxicol Ind Health. 2007;23:411–20.[PubMed]
68. Sypniewska RK, Millenbaugh NJ, Kiel JL, et al. Protein changes in macrophages induced by plasma from rats exposed to 35 GHz millimeter waves. Bioelectromagnetics. 2010;31:656–63. [PubMed]
69. Grigoriev YG, Mikhailov VF, Ivanov AA, et al. Autoimmune processes after long-term low-level exposure to electromagnetic fields part 4. Oxidative intracellular stress response to the long-term rat exposure to nonthermal RF EMF. Biophysics. 2010;55:1054–8.
70. Erdal N, Gürgül S, Tamer L, et al. Effects of long-term exposure of extremely low frequency magnetic field on oxidative/nitrosative stress in rat liver. J Radiat Res. 2008;49:181–7. [PubMed]
71. Ahuja YR, Vijayashree B, Saran R, et al. In vitro effects of low-level, low-frequency electromagnetic fields on DNA damage in human leucocytes by comet assay. Indian J Biochem Biophys. 1999;36:318–22.[PubMed]
72. Amara S, Douki T, Ravanat JL, et al. Influence of a static magnetic field (250 mT) on the antioxidant response and DNA integrity in THP1 cells. Phys Med Biol. 2007;52:889–98. [PubMed]
73. Focke F, Schuermann D, Kuster N, et al. DNA fragmentation in human fibroblasts under extremely low frequency electromagnetic field exposure. Mutat Res. 2010;683:74–83. [PubMed]
74. Franzellitti S, Valbonesi P, Ciancaglini N, et al. Transient DNA damage induced by high-frequency electromagnetic fields (GSM 1.8 GHz) in the human trophoblast HTR-8/SVneo cell line evaluated with the alkaline comet assay. Mutat Res. 2010;683:35–42. [PubMed]
75. Garaj-Vrhovac V, Gajski G, Pažanin S, et al. Assessment of cytogenetic damage and oxidative stress in personnel occupationally exposed to the pulsed microwave radiation of marine radar equipment. Int J Hyg Environ Health. 2011;214:59–65. [PubMed]
76. Hong R, Zhang Y, Liu Y, et al. Effects of extremely low frequency electromagnetic fields on DNA of testicular cells and sperm chromatin structure in mice. Zhonghua Lao Dong Wei Sheng Zhi Ye Bing Za Zhi.2005;23:414–7. [Article in Chinese] [PubMed]
77. Ivancsits S, Diem E, Pilger A, et al. Induction of DNA strand breaks by intermittent exposure to extremely-low-frequency electromagnetic fields in human diploid fibroblasts. Mutat Res. 2002;519:1–13.[PubMed]
78. Ivancsits S, Diem E, Jahn O, et al. Intermittent extremely low frequency electromagnetic fields cause DNA damage in a dose-dependent way. Int Arch Occup Environ Health. 2003;76:431–6. [PubMed]
79. Ivancsits S, Pilger A, Diem E, et al. Cell type-specific genotoxic effects of intermittent extremely low-frequency electromagnetic fields. Mutat Res. 2005;583:184–8. [PubMed]
80. Kesari KK, Behari J, Kumar S. Mutagenic response of 2.45 GHz radiation exposure on rat brain. Int J Radiat Biol. 2010;86:334–43. [PubMed]
81. Lai H, Singh NP. Melatonin and a spin-trap compound block radiofrequency electromagnetic radiation-induced DNA strand breaks in rat brain cells. Bioelectromagnetics. 1997;18:446–54. [PubMed]
82. Lai H, Singh NP. Magnetic-field-induced DNA strand breaks in brain cells of the rat. Environ Health Perspect. 2004;112:687–94. [PMC free article] [PubMed]
83. Lee JW, Kim MS, Kim YJ, et al. Genotoxic effects of 3 T magnetic resonance imaging in cultured human lymphocytes. Bioelectromagnetics. 2011;32:535–42. [PubMed]
84. Paulraj R, Behari J. Single strand DNA breaks in rat brain cells exposed to microwave radiation. Mutat Res. 2006;596:76–80. [PubMed]
85. Romeo S, Zeni L, Sarti M, et al. DNA electrophoretic migration patterns change after exposure of Jurkat cells to a single intense nanosecond electric pulse. PLoS ONE. 2011;6:e28419. [PMC free article] [PubMed]
86. Schwarz C, Kratochvil E, Pilger A, et al. Radiofrequency electromagnetic fields (UMTS, 1,950 MHz) induce genotoxic effects in vitro in human fibroblasts but not in lymphocytes. Int Arch Occup Environ Health. 2008;81:755–67. [PubMed]
87. Svedenstål BM, Johanson KJ, Mattsson MO, et al. DNA damage, cell kinetics and ODC activities studied in CBA mice exposed to electromagnetic fields generated by transmission lines. In Vivo.1999;13:507–13. [PubMed]
88. Svedenstål BM, Johanson KJ, Mild KH. DNA damage induced in brain cells of CBA mice exposed to magnetic fields. In Vivo. 1999;13:551–2. [PubMed]
89. Trosić I, Pavicić I, Milković-Kraus S, et al. Effect of electromagnetic radiofrequency radiation on the rats’ brain, liver and kidney cells measured by comet assay. Coll Antropol. 2011;35:1259–64. [PubMed]
90. Burdak-Rothkamm S, Rothkamm K, Folkard M, et al. DNA and chromosomal damage in response to intermittent extremely low-frequency magnetic fields. Mutat Res. 2009;672:82–9. [PubMed]
91. Fairbairn DW, O'Neill KL. The effect of electromagnetic field exposure on the formation of DNA single strand breaks in human cells. Cell Mol Biol (Noisy-le-grand) 1994;40:561–7. [PubMed]
92. Fiorani M, Cantoni O, Sestili P, et al. Electric and/or magnetic field effects on DNA structure and function in cultured human cells. Mutat Res. 1992;282:25–9. [PubMed]
93. Malyapa RS, Ahern EW, Straube WL, et al. Measurement of DNA damage after exposure to 2450 MHz electromagnetic radiation. Radiat Res. 1997;148:608–17. [PubMed]
94. McNamee JP, Bellier PV, Chauhan V, et al. Evaluating DNA damage in rodent brain after acute 60 Hz magnetic-field exposure. Radiat Res. 2005;164:791–7. [PubMed]
95. Scarfí MR, Sannino A, Perrotta A, et al. Evaluation of genotoxic effects in human fibroblasts after intermittent exposure to 50 Hz electromagnetic fields: a confirmatory study. Radiat Res. 2005;164:270–6.[PubMed]
96. Stronati L, Testa A, Villani P, et al. Absence of genotoxicity in human blood cells exposed to 50 Hz magnetic fields as assessed by comet assay, chromosome aberration, micronucleus, and sister chromatid exchange analyses. Bioelectromagnetics. 2004;25:41–8. [PubMed]
97. Testa A, Cordelli E, Stronati L, et al. Evaluation of genotoxic effect of low level 50 Hz magnetic fields on human blood cells using different cytogenetic assays. Bioelectromagnetics. 2004;25:613–9. [PubMed]
98. Szabó G, Bährle S. Role of nitrosative stress and poly(ADP-ribose) polymerase activation in myocardial reperfusion injury. Curr Vasc Pharmacol. 2005;3:215–20. [PubMed]
99. Moon HK, Yang ES, Park JW. Protection of peroxynitrite-induced DNA damage by dietary antioxidants. Arch Pharm Res. 2006;29:213–7. [PubMed]
100. Sakihama Y, Maeda M, Hashimoto M, et al. Beetroot betalain inhibits peroxynitrite-mediated tyrosine nitration and DNA strand damage. Free Radic Res. 2012;46:93–9. [PubMed]
101. Hybertson BM, Gao B, Bose SK, et al. Oxidative stress in health and disease: the therapeutic potential of Nrf2 activation. Mol Aspects Med. 2011;32:234–46. [PubMed]
102. Ridley AJ, Paterson HF, Johnston CL, et al. The small GTP-binding protein rac regulates growth factor-induced membrane ruffling. Cell. 1992;70:401–10. [PubMed]
103. Wennström S, Hawkins P, Cooke F, et al. Activation of phosphoinositide 3-kinase is required for PDGF-stimulated membrane ruffling. Curr Biol. 1994;4:385–93. [PubMed]
104. Joucla S, Yvert B. Modeling of extracellular neural stimulation: from basic understanding to MEA-based applications. J Physiol Paris. 2012;106:146–58. [PubMed]
105. Pashut T, Wolfus S, Friedman A, et al. Mechanisms of magnetic stimulation of central nervous system neurons. PLoS Comput Biol. 2011;7:e1002022. [PMC free article] [PubMed]
106. Fatemi-Ardekani A. Transcranial magnetic stimulation: physics, electrophysiology, and applications.Crit Rev Biomed Eng. 2008;36:375–412. [PubMed]
107. Silva S, Basser PJ, Miranda PC. Elucidating the mechanisms and loci of neuronal excitation by transcranial magnetic stimulation using a finite element model of a cortical sulcus. Clin Neurophysiol.2008;119:2405–13. [PMC free article] [PubMed]
108. Radman T, Ramos RL, Brumberg JC, et al. Role of cortical cell type and morphology in subthreshold and suprathreshold uniform electric field stimulation in vitro. Brain Stimul. 2009;2:215–28.[PMC free article] [PubMed]
109. Minelli TA, Balduzzo M, Milone FF, et al. Modeling cell dynamics under mobile phone radiation. Nonlinear cell dynamics under mobile phone radiation. Nonlinear Dynamics Psychol Life Sci. 2007;11:197–218. [PubMed]
110. Saunders RD, Jefferys JGR. A neurobiological basis for ELF guidelines. Health Phys. 2007;92:596–603. [PubMed]
111. Havas M. Dirty electricity elevates blood sugar among electrically sensitive diabetics and may explain brittle diabetes. Electromagn Biol Med. 2008;27:135–46. [PMC free article] [PubMed]
112. Havas M. Electromagnetic hypersensitivity: biological effects of dirty electricity with emphasis on diabetes and multiple sclerosis. Electromagn Biol Med. 2006;25:259–68. [PubMed]
113. de Vochta F. “Dirty electricity”: what, where, and should we care? J Expo Sci Environ Epidemiol.2010;20:399–405. [PubMed]
114. Friedman J, Kraus S, Hauptman Y, et al. Mechanism of short-term ERK activation by electromagnetic fields at mobile phone frequencies. Biochem J. 2007;405:559–68. [PMC free article] [PubMed]
115. Desai NR, Kesari KK, Agarwal A. Pathophysiology of cell phone radiation: oxidative stress and carcinogenesis with focus on the male reproductive system. Reproduct Biol Endocrinol. 2009;7:114. doi:10.1186/1477-7827-7-114. [PMC free article] [PubMed]
116. Wyatt CN, Weir EK, Peers C. Diphenylamine iodonium blocks K+ and Ca2+ currents in type I cells isolated from the rat carotid body. Neurosci Lett. 1994;172:63–6. [PubMed]
Articles from Journal of Cellular and Molecular Medicine are provided here courtesy of Blackwell Publishing
 RSS Feed
RSS Feed
